カルシウム代謝とその調節
千勝典子:東京大学大学医学部第四内科
松本俊夫:徳島大学大学医学部第一内科
| はじめに | |||
| 1.カルシウムの生体内分布 | |||
| 2.カルシウムの代謝平衡 | |||
| 3.カルシウム代謝調節ホルモンの作用 | |||
| 1)PTH | ①分泌調節 | ②作用 | |
| 2)1,25(OH)2D | ①産生、分泌調節 | ②作用 | |
| 3)カルシトニン(CT)2D | ①分泌調節 | ②作用 | |
| 4.各臓器でのカルシウム代謝の調節 | |||
| 1)腸管のカルシウム吸収の調節 | |||
| 2)腎尿細管カルシウム輸送の調節 | |||
| 3)骨でのカルシウム代謝の調節 | |||
| おわりに | |||
| 参考文献 | |||
はじめに
カルシウムは生体にとって、構造の維持に必須である骨の主要な構成成分である。と同時に、筋肉の収縮、神経興奮性やホルモン分泌、酵素活性の変化などの各種の細胞機能の調節因子として、生体機能の維持および調節に不可欠な役割を担っている。
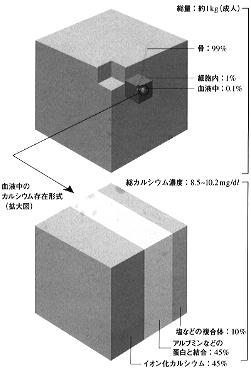
成人の体内には約1Kgのカルシウムが存在する。このうち約99%が骨に、ヒドロキシアパタイト(Ca10(PO4)6(OH)2)の形で存在し、残り約1%のほとんどが細胞内に分布している。血液中には全体の約0,1%(約1g)が存在するにすぎない。血清カルシウムのうち約45%はアルブミンなどの蛋白と結合して、残りの約45%が遊離イオン化カルシウムとして存在する(図1)(*1)。 このうちホルモンによる調節を受け、細胞内外への移行などにより神経、筋肉の興奮性など細胞機能の維持・調節に必須の役割を果たしているのはイオン化カルシウムである。 ヒトをはじめとする陸上動物においては、常に血液中のカルシウム濃度は低下の危機にさらされている。このため血清カルシウムを上昇させる副甲状腺ホルモン(PTH)や活性型ビタミンDである1,25水酸化ビタミンD(1,25(OH)2D)などの働きによりその濃度が維持されている。 カルシウム代謝調節ホルモンの作用発現には、これらのホルモン間の相互作用が重要である。したがって、カルシウム代謝調節系を理解するには、これらのホルモンによる代謝調節のネットワーク全体を把握する必要がある。 |
【図1】
|
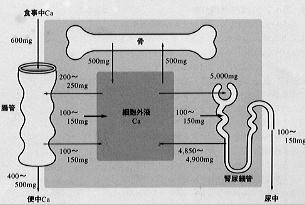
生体を一つのブラックボックスとして考えた場合、腸管からの食物中のカルシウム吸収と、便、尿中へのカルシウム排泄により生体内外のカルシウム代謝平衡が規定される(図2)。
、すなわち、腸管からのカルシウム吸収がカルシウム排泄を上回ればカルシウム・バランスはプラスとなり、逆にカルシウム排泄がカルシウム吸収より多ければカルシウム・バランスはマイナスとなる。
前述のように体内カルシウムの99%以上は骨に存在するため、生体内のカルシウム量の増減はほぼ骨のカルシウム量の変化を示すことになる。したがって、プラスのカルシウム・バランスは骨量の増加を表し、マイナスは骨量の減少を意味する。
【図2】生体のカルシウム |
生理的にも、思春期までの成長期には骨の成長に伴いプラスのカルシウム・バランスが、一方、40歳代以降になると骨量の減少を反映してマイナスのカルシウム・バランスが認められる。 |
生体内外のカルシウムの出入りに加え、生体内の骨と細胞外液、および細胞内と細胞外液との間では一定のカルシウムイオンの濃度勾配が維持されて動的な平衡関係が保たれている。細胞質内のカルシウムイオン濃度は10-8から10-7Mという極めて低い範囲に維持されている。筋肉の収縮、ホルモンの分泌、酵素活性の変化などの細胞機能の維持・調節上必須の役割を演じているのはこの細胞質内カルシウムイオンである。
一方、血清カルシウムイオン濃度は10-3Mの範囲であり、細胞膜を介して大きなカルシウムの濃度勾配が存在している。細胞膜のカルシウム透過性が極めて低いこと、細胞膜には各種のカルシウムポンプが存在することなどによりこのような濃度勾配が維持されている。
血清カルシウム濃度の異常は細胞内カルシウムイオン濃度の維持機構にも変調をもたらし、細胞機能の異常を来しうる。その代表な例が低カルシウム血症のときにみられる痙攣、テタニーや知覚障害であり、高カルシウム血症のときにみられる意識障害、腸管運動障害などである。
、このため血液中のカルシウム濃度は8,5~10,2mg/dlという狭い範囲に常に維持される必要があり、それを調節する副甲状腺ホルモン(PTH)や活性型ビタミンD(1,25(OH)2D)などのカルシウム調節ホルモンである。
、すなわち、カルシウム調節ホルモンは腸管、腎臓、骨などに作用し、これらの臓器と血液との間のカルシウムの出入りを調節することにより血清カルシウム濃度を狭い範囲に維持する役割を果たしている。
副甲状腺より分泌されるPTHはカルシウム代謝調節系の中心をなすホルモンである。副甲状腺からは84個のアミノ酸よりなるポリペプチドとして分泌されるが、肝臓、腎臓などで代謝され、いくつかのフラグメントとなる。生物活性はN端から34個のアミノ酸部分に存在するので、この部分が保たれたフラグメントと分泌されたままの形のPTHが生物作用を発揮する。
PTH分泌を調節するうえで生理的にも最も重要な因子は血清カルシウムイオンである。 |
【図3】副甲状腺ホルモン(PTH)の分泌調節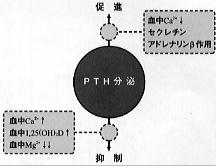 |
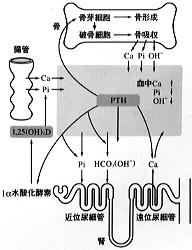
図4に示すようにPTHの主な標的器官は骨と腎臓であるが、腎臓での1,25(OH)2D産生促進作用を介して、間接的に腸管カルシウム吸収も促進される。
 【図4】副甲状腺ホルモン(PTH)の作用 |
PTHは骨に作用し骨吸収を促進する。骨吸収に関わっているのは破骨細胞と呼ばれる多核巨細胞である。ところがPTH受容体は破骨細胞にはなく、骨形成に携わる骨芽細胞に存在する。 この細胞間相互作用に関わる因子として破骨細胞分化因子(ODF)が同定されており、PTHによりその発現が増加することも示されている。 PTHは同時に骨形成も促進し、結果として骨の代謝回転を高める。総じて骨吸収の促進作用が上回るためヒドロキシアパタイト結晶の溶出によりカルシウムイオン、リン(P)、水酸イオン(OH-)が血中に動員される。 |
腎臓におけるPTH作用は多岐にわたる。近位尿細管ではリンパおよびHCO3-の再吸収を抑制することにより、その排泄を促進する、またcAMPの産生を介して1α水酸化酵素を活性化し、1,25(OH)2Dの産生を高める。遠位尿細管ではカルシウム再吸収を促進し血清カルシウム濃度の維持に重要な役割を果たしている。後に述べるご特この作用の発現にはビタミンDの存在が必要である。
これらのPTH作用の総和として、血清カルシウムイオン濃度は上昇するが、骨より動員されたイオンのうちリンおよび水酸イオンの腎臓からの排泄が促進され血中濃度は低下する。
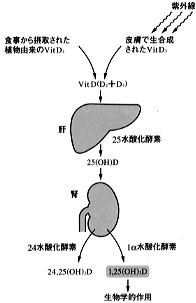
【図5】ビタミンDの代謝(活性型ビタミンDの生合成過程)→ (図5)に示すように、ビタミンDは紫外線の働きによって皮膚においてコレステロール前駆体から生合成されるビタミンD3と、食物にのみ含まれるビタミンD2からなるが、両方ともその代謝経路、作用の点ではほとんど同じである。 ビタミンDは下部小腸から吸収され、ビタミンD結合蛋白と結合して血中を運ばれ、まず肝臓において25位が水酸化されて、25(OH)Dへと変換される。 |
 |
①産生、分泌調節(図6)
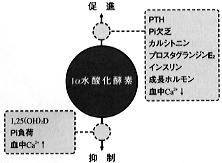 【図6】1,25(OH)2Dの産生調節 |
1,25(OH)2Dの産生における律速段階として最も重要なのは腎臓の1α水酸化酵素であり、この酵素活性は図6のような因子により調節を受けている(*4)。 このうち最も強力な促進因子はPTHである。血清リンの低下も促進因子となる。逆に1,25(OH)2Dは自ら1α水酸化酵素の活性を抑制する。このほか、血清カルシウムイオン、インスリン、成長ホルモン等も1,25(OH)2Dの産生に影響を与える。 |
1,25(OH)2Dは副甲状腺に働いてPTHの合成、分泌を抑制する。また腸管ではカルシウム、リンの吸収を促進する。腎臓においては前述のごとく自らの生成に携わる1α水酸化酵素活性を抑制すると同時に24水酸化酵素の発現を高めることにより25(OH)Dの24,25(OH)2Dへの変換を促進する。この結果、産生される24,25(OH)2Dもいくつかの特異作用を有することが最近明らかになってきている。また、1,25(OH)2Dは遠位尿細管でのカルシウム再吸収を自ら高めるとともに、PTHのカルシウム再吸収作用を増強する。 ビタミンDは骨形成に必須であるが、1,25(OH)2Dの骨形成に対する直接作用については、明確でない点が残されている。1,25(OH)2Dは破骨細胞の形成過程にも必要であり、PTHの骨作用の発現に影響を及ぼすとともに、高濃度では自ら骨吸収を促進する。 |
【図7】1,25(OH)2Dの作用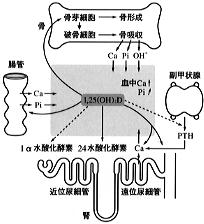 |
CTは32個のアミノ酸よりなるポリペプチドホルモンで甲状腺のC細胞から分泌される。
血清カルシウムイオン濃度の上昇により分泌が促進される。このほか、ガストリン、GIPなどの消化管ホルモンによっても分泌が促進される。
骨において、CTは破骨細胞に作用することにより骨吸収を抑制する(*5)。CTの受容体は破骨細胞にしか存在しないが、骨吸収の抑制により二次的に骨芽細胞による骨形成も抑制される。CTの骨吸収抑制作用はCTの長期投与により減弱する(エスケープ現象)。
CTは腎でのカルシウム、リンの再吸収にも影響を及ぼすが、この作用には種差が大きく、ヒトでの作用は必ずしも明確ではない。CTはまた、尿細管の1α水酸化酵素活性を促進することにより1,25(OH)2Dの産生を高める。この作用はPTHと異なる部位で(近位直尿細管)、異なる作用機序により発現する考えられている。
わが国では成人は1日600mgのカルシウムを摂取するといわれている。このうち200mgあまりが腸管から吸収されるが、消化液の分泌などに伴い体内から消化管中に100mg程度が分泌されるため正味100~150mgが体内に入ることになる。カルシウムの吸収は十二指腸などの上部腸管で主に営まれているが、空腸、回腸でも行われる。腸管からのカルシウム吸収を調節するうえで最も重要なホルモンは活性型ビタミンD(1,25(OH)2D)である。
1,25(OH)2Dは腸管からのカルシウムの吸収効率を高めるが、血清1,25(OH)2D濃度は生体のカルシウム需要に応じてPTH分泌などを介して変化する。こうして食事中のカルシウムがある程度変化しても、吸収されるカルシウム量は必要に応じて調節されている。しかし、カルシウムの摂取量が大幅に低下すると吸収量も不足することになる。一方、食事中のカルシウム量が著しく高まると細胞間を通過する受動的なカルシウム吸収が増加し、高カルシウム尿症や高カルシウム血症がもたらされうる。
1,25(OH)2Dの腸管での作用機序については、今なお不明な点が残されている。一般にステロイドホルモンは、核内に存在するレセプターに結合し特定の蛋白の転写、翻訳を調節することによりその作用を発現すると考えられてる。腸管上皮細胞においても、1,25(OH)2Dがカルシウム結合蛋白の合成を誘導することから、このカルシウム結合蛋白を介してカルシウム吸収を促進する可能性が考えられる。しかしながら、カルシウム結合蛋白がカルシウム吸収に直接関与することを示す成績は得られていない。
一方、1,25(OH)2Dは腸管上皮細胞膜のリン脂質構造を変化させることから、これがカルシウム吸収の促進に関与している可能性がある。また、ニワトリ腸管灌流系では1,25(OH)2D投与後十数分でカルシウム輸送が促進されることから、1,25(OH)2Dが受容体を介さず直接細胞膜に作用している可能性も考えられている。1,25(OH)2Dの腸管カルシウム吸収作用の発現機序については今後の検討が必要である。
腎糸球体からは血液中の蛋白などと結合していない遊離カルシウム(約5mg/dl)が濾過され、GFRが100㍑/日の人ではその量は1日約5,000mgに達する。しかし、濾過されたカルシウムの50~55%は近位尿細管で、20~30%がヘンレ係蹄、10~15%が遠位尿細管で、さらに2~8%が集合管で再吸収されると考えられている。したがって、糸球体から濾過されたカルシウムの95%以上が腎尿細管において再吸収を受けることになる。
、このうち近位尿細管でのカルシウム再吸収は、水、ナトリウム(Na)の輸送に伴う受動的なものであり、ヘンレ上行脚でも塩素イオンの輸送により生じる電位差に依存する二次的なカルシウム輸送の依存が確認されている。
、これに対し遠位尿細管でのカルシウム輸送はナトリウム再吸収や電位差に依存しておらず、またPTHの作用により調節を受ける。したがって量的には近位尿細管におけるカルシウム再吸収が多いもののホルモンによる調節を受け、最終的な尿中カルシウム排泄量を決定する部位は遠位尿細管である。通常、成人では1日100~150mgの尿中カルシウム排泄が認められる。
PTHがどのような機序でカルシウム再吸収を促進するかについても不明な点が多い。PTHは標的細胞に作用し、アデニレートシクラーゼを活性化することによりサイクリックAMP(cAMP)の産生を亢進させ、このcAMPをセカンドメッセンジャーとして多くの作用を発現する。腎臓においても、近位尿細管ではcAMP産生の促進を介して25(OH)Dから1,25(OH)2Dへの変換などを促進する。腎遠位尿細管にもPTH反応性アデニレートシクラーゼが存在することから、従来PTHによるカルシウム再吸収促進作用もcAMPを介して発現する可能性が考えられていた。しかし、現在は遠位尿細管におけるPTHのカルシウム再吸収促進作用については、活性型ビタミンDの存在下でcAMPを介さないメカニズムにより発現しうるものと考えられている。
尿中カルシウム排泄はPTH、1,25(OH)2Dのほかにも図8に示す種々の因子により影響を受ける。
脱水、ナトリウム負荷は近位尿細管でのナトリウム、水の再吸収を介してカルシウム再吸収をそれぞれ促進、あるいは抑制する。 フロセマイドはヘンレ上行脚での塩素イオンの輸送を抑制することにより、電位差依存性のカルシウム再吸収を抑制する。一方、サイアザイドは主に遠位尿細管でのカルシウム再吸収を促進する。したがって、フロセマイドが高カルシウム尿症の高カルシウム尿症の治療薬として用いられのに対して、サイアザイドは高カルシウム血症の患者には禁忌であり、腎型の高カルシウム尿症患者の治療に使用される。 |
【図8】腎尿細管カルシウム再吸収に影響する因子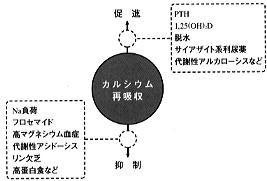 |
マグネシウム(Mg)はカルシウムと同じく主にヘンレ上行脚で再吸収されるため、高マグネシウム血症、高マグネシウム尿症ではマグネシウムがカルシウムと競合することによりカルシウムの再吸収が阻害される。
骨は破骨細胞による骨吸収と骨芽細胞による骨形成を常に繰り返す動的組織である(骨のリモデリング)。
健常な成人では破骨細胞による骨吸収と骨芽細胞による骨形成量はほぼ等しく、骨吸収と骨形成の両過程間には共役関係(カップリング)がある。すなわちPTHや1,25(OH)2Dなどの骨吸収を促進する因子は、骨表面の骨芽細胞にまず作用し、これを介して破骨細胞により骨が吸収される。 |
【図9】骨のリモデリング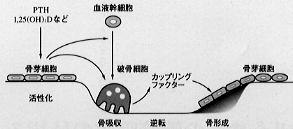 |
骨吸収によりヒドロキシアパタイト結晶が融解されると、カルシウム、リン酸、水酸イオンが血液中に放出される。一方、破骨細胞から分泌される蛋白分解酵素の作用などにより骨基質蛋白も溶出されるが、この中には骨形成に関与するサイトカインも含まれている。これらサイトカインの作用などにより、骨吸収を終えた部位に新たに骨が形成される。
このように骨芽細胞と破骨細胞は機能上、密接に結びついており、両者の連関により、すなわち、骨形成と骨吸収のバランスが保たれる結果、骨量およびカルシウム代謝平衡が維持されている。これに対し骨形成と骨吸収のバランスが崩れると(アンカップリング)、骨量の増加、あるいは減少が生じる。前述した成長期の骨量の増加、壮年期以降の骨量の減少という生理的変化に加え、骨粗鬆症などの骨代謝疾患による骨量の低下が臨床的には特に問題となる。すなわち、骨吸収に比し十分な骨形成が行われない結果、骨量の減少を来し、骨強度が低下して骨折を起こしやすくなる。
生体内のカルシウム分布およびその役割、カルシウムの代謝動態およびその調節機構についてまとめた。
、細胞がその機能を維持するためには、細胞内外のカルシウムイオン濃度の調節が保たれていることが必須である。陸上生活を営む生体では、カルシウムを蓄積し、体内での需要に応じてこれを動員する貯蔵庫が必要である。
、骨は荷重に抗して生体を支え、その運動機能を維持する支持体としての機能に加え、必要に応じてカルシウムを動員する貯蔵庫としての機能という、二つの相反する役割を担っている。
、したがって、カルシウム摂取の不足やカルシウム調節ホルモン作用の異常は、骨代謝異常ひいては骨の支持体としての機能異常をもたらし、場合によっては細胞内外のカルシウム濃度の変化による生体機能の障害をも来す。
、カルシウム代謝調節系を解明し、これを理解することは、これらの異常に基づく疾患の治療の上からも、また予防の面からも極めて重要であると考えられる。
| (*1) | 松本俊夫 : カルシウム代謝調節系、実験医学 10:510-515,1992 |
| (*2) | 徳丸久、清野桂記 : 副甲状腺生合成の分子生物学、日本臨床 53:811-815,1995 |
| (*3) | Okazaki T et al : Conserved mechanism of negative gene regulation by extracellular calciium Parathyroid homone gene versus atrial natriuretic polypeptide gene .J Clin Invest 89:1268,1992 |
| (*4) | 福本誠二 : 骨,カルシウム代謝調節ホルモン.日内会誌 82:1923-1927,1993 |
| (*5) | 塩井淳、西沢良記 : カルシトニン,腎ト透析 37(2);238-242,1994 |