カルシウム不足と記憶 藤田拓男 |
|
はじめに
カルシウムはすべての細胞の情報伝達機構に決定的に重要な役割を演じ、その作用はきわめて厳密に保たれている血清カルシウム血と、その10,000分の1しかない細胞内遊離カルシウム値との間の大きな落差と、これによって維持される緊張状態によって保たれている。
、脳の細胞もその例外ではなく、記憶のような高度な機能も個々の細胞の伝える情報とその総合的な統合に基づき、細胞のカルシウム代謝が正常に維持されていることがその基本的条件である。
、記憶に特に関係の深いといわれている海馬の細胞には、細胞の内外のカルシウムバランスの崩壊が起こりやすいことが知られている。ここにカルシウムと記憶との接点がある。低酸素血症や脂質過酸化物その他の細胞毒による細胞機能障害とともに、細胞膜をカルシウムが通過しやすくなり、細胞内遊離カルシウが増加し、これによって細胞死に至る最終共通経路が進む(*1,2)。
このような細胞レベルでのカルシウム代謝の異常が、栄養摂取その他の全身的なカルシウム代謝の異常、ことにわが国でも世界のほかの国々と同じように広くみられるカルシウム不足とどのように関連しているかを知ることは、老年に多くみられる記憶障害に対して、栄養、生活様式または薬剤療法等の対策によって、実際どのような対策をとることができるかを見きわめて、またこれを実行に移すうえで重要なことである。
1.加齢と記憶障害、痴呆
加齢とともに低下していくいくつかの機能のなかで、視力の調節とともに、記憶記銘障害は、最も目立つものの一つである。多少とも記憶のおとろえを感じない人は、中年以降ほとんどないと思われるほどである。さらにこれが進行して起こる痴呆は、加齢とともに着実に進行し80歳以上では20%に達する。
、老年期にみられる痴呆には、脳血管障害性のものと、血管障害とこれによる虚血性変化の明らかでないいわゆるアルツハイマー型痴呆があり、後者は正常の加齢変化によると思われる記憶障害と連続的に移行するともいわれる(*1.2)。
、アルツハイマー型痴呆に特有の病理学的変化といわれる老人斑や原線維変化は加齢とともに増加し、記憶力の低下と相関する。加齢とともに起こる脳重量の減少、ニュートロン数の減少、リポフスチン色素の増加は比較的非特異的なものであるが、アルツハイマー原線維変化、老人斑、平野斑等はアルツハイマー型痴呆に特異的である。
、しかしながら、一部はパーキンソン病や筋萎縮性側索硬化症のような神経系の変性疾患でもみられ、両者の類似性が指摘されている(*4,5)。
、脳のカルシウム含量は加齢とともに、またアルツハイマー病やその他の神経変性疾患で増加している。カルシウム摂取の不足と平行して起こったいくつかの神経系変性疾患すなわち紀伊半島の筋萎縮性側索硬化症、グアム島のパーキンソン痴呆症候群で腎不全や加齢とともにみられる神経系のカルシウム含量の増加と同様の変化がみられることは、共通の原因を示唆して興味深い。
| 、すなわちカルシウムの不足は続発性副甲状腺機能亢進を起こし、副甲状腺ホルモンが骨からカルシウムを動員するとともに、脳の細胞の中にカルシウムを入れて細胞内外のカルシウムのバランスをくずし、その機能を低下させることが考えられる。アルツハイマー型痴呆は女性では男性より多いが、腸管からのカルシウムの吸収を障害し、骨粗鬆症の原因として知られるエストロゲンの欠乏も、その危険因子の一つであるとされる(*6)(図1)。 | 【図1】脳細胞のカルシウム代謝の調節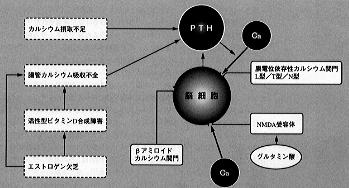 |
2.海馬細胞とカルシウム
アルツハイマー型痴呆の危険因子の一つに脳外傷があるが、外傷・虚血等脳組織内での異常な出来事のあったとき、海馬が低酸素状態に弱いことは古くから知られている。虚血による細胞死は非常に速く、数時間で起こるとされていたが、これより遅く、数日後までに海馬のCAIの細胞死が起こることがわかり遅発性神経細胞壊死と呼ばれている(*7)。これに先立って神経細胞内の遊離カルシウムが増加する(*8)。プログラムされた細胞死すなわちアポトーシスともこの点に共通点がある(*9)。
、活性型アミノ酸であるグルタミン酸はNMDA受容体を介して神経細胞内へのカルシウムの流入を促進することが知られているが、海馬領域で特にグルタミン酸の産生が多いとはいえないようである。
、記憶はシナプスの可塑性の結果起こるもので特定の回路が強化されて残ることの発現と考えられるが、海馬のシナプスには非常に強い可塑性がある。この結果、強い刺激を与えると長期増強現象がみられる。細胞内遊離カルシウムの増加によってこの現象が起こることは明らかであるが、NMDA受容体以外にもアセチルコリンが複雑に関与していることがわかった。ムスカリン作動性のアトロピンで抑制される機序で、低量のアセチルコリンは細胞内遊離カルシウムを上昇させるが、ピレンゼピンによるMI遮断の後には細胞内遊離カルシウムをかえって低下させる作用がある。これらの作用のなかにはグリア細胞を介して起こると思われるものもあり、グリア細胞にはアセチルコリンに反応するものが多い。
3.脳細胞のカルシウム代謝
アルツハイマー型痴呆を示す患者の脳では、海馬のほかに大脳皮質前頭葉や側頭葉でもカルシウム含量が増加するが、このときNa+/Ca2+の交感が促進していることが認められた(*9,10)。カルシウム摂取不足のときに必ず分泌が亢進し、その全身的なカルシウム代謝の影響ことに骨からのカルシウムの流出と脳その他の軟部組織への流入、細胞外から細胞内へのカルシウムの流入を通して記憶その他の脳の機能や痴呆の発現にも影響を与える。
、副甲状腺ホルモンは進化的には比較的新しいホルモンであり、カルシウムの豊富な海水中に住む魚類では認められず、カルシウムの欠乏する陸上の環境に両棲類以降にはじめてみられるが、副甲状腺だけから産生されると考えられてきた。しかしながら脳ことに視床下部や下垂体からも副甲状腺ホルモンと区別しがたい物質が分泌されるようである(*11)。
、PTHのmRNAは視床下部の傍室核と視束上核に局在し、この部位における合成が確認された(*12)。分泌刺激としては副甲状腺におけると同様に、低カルシウム血症やカルシウムの欠乏が脳からのPTHの分泌を起こすと思われる。副甲状腺ホルモン関連ペプチド(PTHrP)はPTHと異なり血管等を含むいろいろな組織から分泌され、パラクリン的に作用すると考えられているが、脳でも産生が認められている。
、魚類のスタニウス体で産生されるPTHと似たペプチドホルモンであるスタニオカルシンはPTHと同様の脳内分布を示す。腎不全の続発性副甲状腺機能亢進で認められる透析脳症すなわち意識混濁、譫妄等は、血清カルシウムレベルに関係なくPTH投与で再現することができ、またPTHは可塑的に脳波の徐波化を起こし、大量では中枢神経系に対して有害な作用をもつものと思われる(*13)。
、森本らは血中PTHと老人の認識反応との間に負の相関を認めた(*14)。脳室内にPTHを注射すると学習・記憶能力の低下が認められる(*15)。痛覚はPTHによって亢進することが認められた(*16)。PTHはL型のカルシウム関門を刺激して神経細胞内へのカルシウムの流入を促進する(*17)。ラット脳のシナプトソームからのノルアドレナリンの放出はPTHによって抑制され、またNaKATPase活性の抑制を介してその取り込みも減少させる(*18)。ラット視床下部のドパミン含量もPTHによって減少する。PTHの脳細胞内カルシウム上昇作用とドパミン減少作用はcAMPを介するものではないと思われる。神経系および脳にPTHの特異的な受容体があるとされる。
カルシウムの不足に影響されるのは神経細胞よりも、むしろ星状細胞またはグリア細胞かもしれない。星状細胞はグルタミン酸に対して細胞内遊離カルシウムの増加で反応するが、最初のスパイク状の上昇はメタボトロピックの機序でIP3に関係し、その後の持続的な上昇はイオノトロピックな機序によると考えられる。これは細胞からの膜電位依存性のカルシウム関門を介しての流入によると思われる(*19)。グリア細胞とニュートロン細胞の間には相互作用があり、ニュートロン細胞なしでグリア細胞だけを培養すると、時折細胞内遊離カルシウム濃度のスパイク様上昇がみられるだけであったが、ニュートロン細胞と一緒に培養するとカルシウム波が発生伝播する。ニュートロン細胞に対して機械的刺激が加えられると、これに接するグリア細胞にもカルシウム波が発生し、両者の接触による密接な機能的関係がうかがわれる(*20)。
カルシウム・パラドックスとは再灌流障害ともいわれる現象であって、心筋細胞をカルシウムを含まない液で灌流した後、今度はカルシウムを含む液で灌流すると、細胞機能が障害され、細胞死に至るものである。このようなカルシウム欠乏のあとに起こるカルシウムの細胞障害作用はグリア細胞にもみられ、ナトリウムとカルシウムの交感機序によるとされる(*21)。全身的なカルシウムの欠乏が副甲状腺ホルモンの分泌を介して細胞内のカルシウムの増加につながることも、老化のカルシウムパラドックスといわれる。
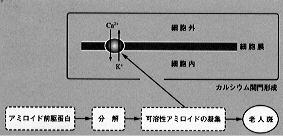
アルツハイマー型痴呆の患者の脳にみられるβアミロイドは細胞膜に存在する蛋白の一つであって、カルシウム関門として働き、細胞内の遊離カルシウム濃度を上昇させることによって細胞の機能障害と死を起こすことが認められている(*22)(図2)。 |
【図2】βアミロイド蛋白 - カルシウム関門説
 |
カルシウムと骨代謝には日内変動があり、夜間は血清カルシウムはやや低下し、副甲状腺ホルモンの分泌が増加することが知られており、このとき骨の吸収も増加する。アルツハイマー病患者では夕方から夜にかけて不穏状態が起こり、たそがれ症候群として知られているが、血清カルシウムの低下が正常者より顕著であり、副甲状腺ホルモンも高く径口的に大量の吸収のよいカルシウムを投与すると血清カルシウムの低下を止め症状を改善する場合がある(*23)。
おわりに
カルシウムは神経細胞の情報伝達にもきわめて重要な役割を果たし、カルシウムの欠乏による副甲状腺ホルモンの分泌の亢進は、脳ことに記憶に重要であるとされる海馬細胞で細胞内遊離カルシウムを増加させることによってその機能を障害し、細胞死に至らせ痴呆の原因となる可能性がある。夕方から夜にかけて血清カルシウムが低下し副甲状腺ホルモンの分泌が亢進することは、アルツハイマー病患者にみられる不穏状態や症状の増悪と関係があるかもしれない。
文 献
| (*1) | Chen ST, Hsu CY, Hogan EL, Juan MS, Banik NL, Balentine JD : Brain calcium content in ischemic infarction. Neurobiol 37:1227-1229,1987 |
| (*2) | Siesjo BK : Calcium and ischemic brain damage. Eur Neurol 25(Suppl1):45-56,1986 |
| (*3) | Brayne C, Calloway P : Normal aging, impaired cognitie function and senile dementia of the Alzheimer type. A continuitum? Lancet 1:1265-1267,1988 |
| (*4) | Yase Y : Calcium, metals and nervous system in the elderly. J Neurol Sci Vitaminol 31(Suppl):S37-S40,1985 |
| (*5) | Wilcock GK : Alzheimer's disease. Current issues. Quart J Med (New Series 66)250:117-124,1988 |
| (*6) | Paganini-Hill A, Henderson VW : Estrogen deficiecy and risk of Alzheimer's disease in women. Am J Epidemiol 140:256-261,1994 |
| (*7) | Nakanura T, Ozawa A, Kudo Y : Calcium-dependent neuronal cell death. Mol Neurol Biol Brain Ischemia. Springer,75-94,1996 |
| (*8) | 工藤佳久:海馬ニュートロンにおけるCaの動態、神経研究の進歩 38:103-111,1994 |
| (*9) | Schwartzman RA, Cidiowski JA : Apoptosis. The biochemistry and molecular biology of programmed cell death 14:133-151,1993 |
| (*10) | Colvin RA, Davis N, Wu A, Murphy CA, Levengood J : Studies of the mechanism underlying in creased Na+/Ca2+exchange activity in Alzheimer's disease brain. Brain Res 665:192-200,1994 |
| (*11) | Balabanova S, Tollner U, Richter HP, Pohland TF, Gaedicke G, Teller WM : Immunoreactive parathyroid hormone, calcium and magnesium in human cerebrospinal fluid. Acta endocrinol 106:227-233,1984 |
| (*12) | Harver S, Frase RA : Parathyroidhormone ; Neural and neuroendocrine perspectives. J Edocrinol 139:353-361,1993 |
| (*13) | Arieff AI, Massry SG : Calcium metabolism of brain in acute renal failure. J Clin Invst 63:387-392,1974 |
| (*14) | Morimoto S, Masugi F, Hironaka T, Shiraishi T, Hoh K, Yamamoto H, Koh E, Onishi T, Ogihara T, Kumahara Y : Relation of serum parathyroid hormone to cognitive function in elderiy females. New Actions of Parathyroid Hormone.329-334,SG Massrt, T Fujita eds. Plenum 1988 |
| (*15) | Clementi G, Drago F, Prato A, Cavaliere S, Rodolico G : Effects of calcitonin, parathyroid hormone and its related fragments on aquisition of active avoidance behavior. Physiol Behavior 33:913-916,1984 |
| (*16) | Gennari C, Agnusdei D, Gonneli S, Conte F, Camporeali A, Palamieri R, Zacchei F : Hyperalgic activity of parathyroid hormone. J Endocrinol Invest 14(Suppl3):34,1991 |
| (*17) | Dubovsky LS, Franks RD : Intracellular calcium ions in affective disorders : Areview and a hypothesis. Biol Psychiat 18:781-797,1983 |
| (*18) | Massry SG, Smogorzewski M : Derrangement in brain synaptosome function in chronic renal failure : Role of parathyroid hormone. New Actions of parathyroid hormone.301-316 SG Massry and T Fujita eds. Plenum,1988 |
| (*19) | Kim WT , Rioult MG, Cornelbell AH : Glutamate-induced calcium sign in astrocytes. Glia 11:173-184,1994 |
| (*20) | Charles AC : Glia-neuron intercellular calcium signaling. Development Neursci 16:196-206,1994 |
| (*21) | Kimlee MH, Stokes BT, Yates AJ : Reperfusion paradox - A new mode of glia cell injury. Glia 5:56-64,1992 |
| (*22) | Arispe N, Oollard HB, Rojas E : β-amyloid Ca2+-channel hypothesis for neural death in Alzheimer disease. Mol Cell Biol 140:119-125,1994 |
| (*23) | Fujita T, Fujita Y, ohgitani S, Iwamoto B, Miki K, Takahashi Y, Matsuoka H, Ando S, Morita Y : Exaggerated evening fall of serum Ca as a possible factor in sun-down syndrome in Alzheimer's disease. Fifth intern Cont on Alzheimer's disease and Related Disorders Abs 681,1996 |